анатомия коры головного мозга
Анатомия коры головного мозга
Кора представлена слоем серого вещества толщиной 3-5 мм. В коре насчитывают до 15 и более млрд. нейронов, а число глиоцитов в головном мозге — более 100 млрд.
Развитие. Формирование коры больших полушарий происходит путем закономерной миграции нейробластов эпендимного слоя вдоль вертикально ориентированных радиальных глиоцитов. Первыми возникают наиболее поверхностный и глубокий слои коры. Затем возникают очередные последовательные волны миграции групп нейробластов, которые дифференцируются в нейроны V-гo, затем IV-гo слоя и д. т. Таким образом, нейробласты очередной волны миграции преодолевают слой нейронов, возникших от более ранней волны миграции. Это создает послойную (экранную) цитоархитектонику коры большого мозга.
Между нейронами устанавливаются сложные взаимосвязи в соответствии с их местом в составе рефлекторных дуг. Формируются ядерные и экранные нервные центры. Тесные взаимоотношения в ходе гистогенеза складываются между нейронами и клетками глии.
Строение. Все нейроны коры мулътиполярные. Среди них по форме клеток различают пирамидные и непирамидные (звездчатые, корзинчатые, веретенообразные, паукообразные и горизонтальные) нейроны. Пирамидные нейроны, наиболее характерны для коры, имеют тело, по форме напоминающее пирамиду, вершина которой обращена к поверхности коры.
От основания пирамидной клетки отходит аксон, имеющий коллатерали (возвратные, горизонтальные, косые). От вершины и боковых поверхностей тела отходят длинные дендриты (апикальные и базальные). Апикальные дендриты группы нейронов объединяются в дендритические пучки. На поверхности дендритов одного пирамидного нейрона может быть до 4-6 тыс. специальных рецепторных аппаратов — шипиков. Наличие в последних актомиозинового комплекса позволяет изменять площадь синаптического контакта, следовательно, влиять на синаптическую связь.

Величина тела пирамидных клеток варьирует от 10 до 150 мкм. Различают малые, средние, крупные и гигантские пирамиды. Пирамидные клетки — это эфферентные нейроны коры, их аксонные коллатерали образуют 3/4 всех синапсов в коре.
Звездчатые нейроны имеют тело в форме звезды. Дендриты отходят во все стороны от тела звездчатого нейрона. Они в большинстве случаев короткие и лишены шипиков. Аксоны звездчатых клеток образуют сложные разветвления около клетки. Это так называемые околоклеточные паутинообразные аксонные сети. Эти клетки встречаются в нижних слоях коры.
Корзинчатые клетки (мелкие и крупные), расположены во П-м и Ш-м слоях коры, своими многочисленными отростками образуют синаптические связи с телами пирамидных нейронов V-ro слоя. Клетки содержат медиатор (ГАМК), который тормозит передачу возбуждения.
Нейроглиоморфные клетки встречаются во всех слоях коры. Это мелкие мультиполярные нейроны с короткими ветвящимися денд-ритами и аксонами.
Биполярные нейроны — малочисленная группа клеток, от тела которых отходят аксон и дендрит. В целом соотношение между пирамидными и другими формами нейронов равно 85:15, то есть в пользу пирамидных нейронов.
Цитоархитектоника. В двигательной зоне коры различают шесть основных слоев: молекулярный, наружный зернистый, пирамидный, внутренний зернистый, ганглионарный, слой полиморфных клеток.
В первом (наружном) молекулярном слое почти нет тел нейронов. Обнаруживаются единичные горизонтально ориентированные нейроны, тангенциальные ветвления нервных волокон нижележащих нейронов и глиальные клетки.
Второй, или наружный зернистый, слой содержит мелкие звездчатые и пирамидные нейроны размером около 10 мкм. Аксоны этих нейронов оканчиваются в III-м, IV-м и VI-м слоях коры, а дендриты поднимаются в молекулярный слой.
Третий слой — это слой средних и крупных пирамидных нейронов. Аксоны этих клеток образуют ассоциативные нервные волокна, идущие через белое вещество и соединяющие соседние участки коры.
Четвертый, или внутренний зернистый, слой содержит в основном мелкие звездчатые нейроны. Аксоны этих клеток, разветвляются в пределах соседних как выше, так и ниже лежащих слоев коры. Этот слой сильно развит в зрительной и слуховой зонах коры. Он состоит из сенсорных звездчатых нейронов, имеющих многочисленные ассоциативные связи с нейронами других типов.
Пятый — ганглионарный — слой образован большими пирамидными нейронами (клетками Беца). Апикальные дендриты нейронов направляются в молекулярный слой. Аксоны этих клеток идут в белое вещество, образуя комиссуральные и проекционные нервные волокна, и прежде всего пирамидные пути.
Шестой слой — слой полиморфных нейронов — содержит также много эфферентных пирамидных нейронов. Кроме того, здесь есть веретенообразные нейроны. Дендриты нейронов шестого слоя пронизывают всю толщину коры, достигая молекулярного слоя.
Анатомия коры головного мозга
а) Доли мозга. Поверхность коры полушарий головного мозга покрыта бороздами и расположенными между ними складками — извилинами. Большая часть коры полушарий скрыта от обзора в углублениях борозд. Формы борозд у разных людей могут отличаться, однако определенные борозды достаточно схожи и могут служить описательными ориентирами. Самые глубокие—латеральная (сильвиева) и центральная (роландова) борозды.
Эти борозды разделяют полушария на четыре доли (в боковой проекции) двумя воображаемыми линиями. Первая линия проходит от латеральной борозды назад, вторая — от верхнего края теменно-затылочной борозды к предзатылочной вырезке, расположенной в нижней границе полушария (борозда и вырезка обозначены на рисунке ниже). Выделяют лобную, теменную, затылочную и височную доли мозга.
Наиболее выступающие участки лобной, затылочной и височной долей образуют полюса полушарий головного мозга.
При удалении краев латеральной борозды (покрышки) открывается доступ к островку головного мозга. В статье по эмбриогенезу головного мозга отмечено, что во время развития конечного мозга плода островок мозга остается относительно «неактивным».
 Пять долей мозга. (А) латеральная и (Б) медиальная поверхности правого полушария мозга.
Пять долей мозга. (А) латеральная и (Б) медиальная поверхности правого полушария мозга.  Правое полушарие с (А) латеральной и (Б) медиальной стороны с изображением основных борозд и извилин.
Правое полушарие с (А) латеральной и (Б) медиальной стороны с изображением основных борозд и извилин.
Обратите внимание на рисунки ниже, а также приведенное ниже описание особенностей поверхности долей мозга.
1. Лобная доля. На латеральной поверхности лобной доли расположена прецентральная извилина, спереди ограниченная прецентральной бороздой. Прямая, верхняя, средняя и нижняя извилины лобной доли ограничены верхней и нижней лобными бороздами. На медиальной поверхности верхняя лобная извилина отделена от поясной извилины поясной бороздой. На нижней (глазничной) поверхности расположены несколько глазничных извилин. Обонятельная луковица и обонятельные пути соприкасаются с глазничной поверхностью лобной доли.
2. Теменная доля. На передней части теменной доли расположена постцентральная извилина, ограниченная сзади постцентральной бороздой. Внутритеменная борозда разделяет заднюю часть теменной доли на верхнюю и нижнюю теменные дольки. В нижней части теменной доли расположены надкраевая извилина, огибающая концевой изгиб латеральной борозды, и угловая извилина, огибающая верхнюю височную борозду.
На медиальной поверхности теменной доли располагается задняя часть околоцентральной дольки мозга, а позади нее — предклинье полушарий. Название околоцентральной дольки, частично расположенной в лобной доле, обусловлено ее положением относительно центральной борозды.
3. Затылочная доля. Латеральная поверхность затылочной доли представлена несколькими боковыми затылочными извилинами. На медиальной поверхности расположена клинообразная извилина (клин), разделяющая теменно-затылочную и шпорную борозды. На нижней поверхности расположены три извилины и три борозды. Латеральная и медиальная затылочно-височные извилины разделены затылочно-височной бороздой. Коллатеральная и шпорная борозды ограничивают язычковую извилину.
4. Височная доля. Височная доля имеет верхнюю, среднюю и нижнюю височную извилины, разделенные верхней и нижней височными бороздами. Нижняя поверхность доли представлена передними участками затылочно-височной извилины. Язычковая извилина переходит в парагиппокампальную извилину, которая оканчивается в области крючка гиппокампа — закругленного выступа, расположенного медиально. Как и показано на рисунках срезов мозга, парагиппокампальная извилина располагается под «закрученным» участком коры — гиппокампом.
5. Лимбическая доля. Пятая, лимбическая, доля мозга расположена на медиальной поверхности полушарий. Поверхность лимбической доли представлена поясной и парагиппокампальной извилинами. Обычно говорят о лимбической системе, в состав которой входят гиппокамп, свод мозга и миндалевидное тело.
 Островок мозга (покрышка удалена).
Островок мозга (покрышка удалена). 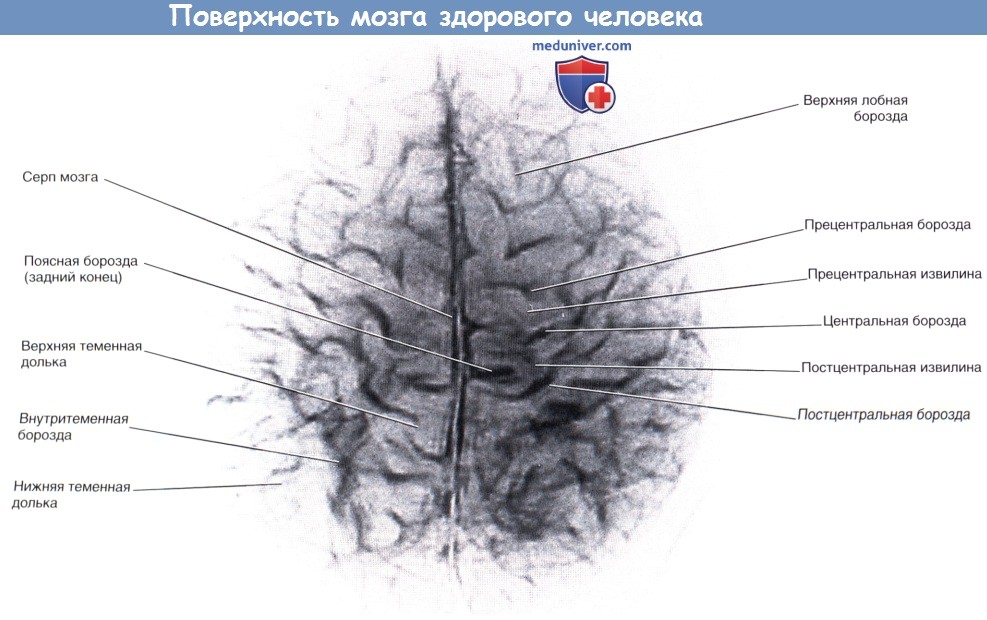 Строение поверхности мозга здорового человека («толстый» МР-срез).
Строение поверхности мозга здорового человека («толстый» МР-срез).  Мозг (вид снизу) с изображением основных борозд и извилин.
Мозг (вид снизу) с изображением основных борозд и извилин.
б) Промежуточный мозг. Основные структуры промежуточного мозга — таламус и гипоталамус. Эти скопления ядер образуют боковые стенки III желудочка. Между таламусом и гипоталамусом проходит гипоталамическая борозда, которая представляет собой ростральную границу пограничной борозды эмбриона.
в) Срединная сагиттальная проекция мозга. Представленный на рисунке ниже срединный сагиттальный срез головы трупа демонстрирует расположение мозга относительно окружающих его структур.
 Промежуточный мозг и его границы. Мозолистое тело.
Промежуточный мозг и его границы. Мозолистое тело.  Сагиттальный МР-срез мозга живого человека.
Сагиттальный МР-срез мозга живого человека.  Сагиттальный срез фиксированного препарата мозга.
Сагиттальный срез фиксированного препарата мозга.
Видео урок анатомия борозд и извилин головного мозга
Редактор: Искандер Милевски. Дата публикации: 9.11.2018
Анатомия коры головного мозга
По иронии судьбы, из всех отделов головного мозга меньше всего мы знаем о функциях коры большого мозга, хотя совершенно очевидно, что это самая большая часть нервной системы. В тоже время известны эффекты поражения или специфической стимуляции различных участков коры. В первых статьях по физиологии на сайте обсуждаются фактические данные о функциях коры; затем кратко представлены основные теории о нервных механизмах, участвующих в мыслительных процессах, памяти, анализе сенсорной информации и др.
а) Физиологическая анатомия коры большого мозга. Функциональном частью коры большого мозга является гонкий слои нейронов, покрывающий поверхность всех извилин. Толщина этого слоя составляет лишь 2-5 мм, а общая площадь равна примерно 0,25 м. В целом кора большого мозга содержит примерно 100 млрд нейронов.
На рисунке ниже показана типичная гистологическая структура коры большого мозга с последовательными слоями разных типов нейронов.
Большинство нейронов относят к трем типам:
(1) зернистые (также называемые звездчатыми);
(3) пирамидные, получившие свое название за характерную пирамидальную форму.
Зернистые нейроны обычно имеют короткие аксоны и, следовательно, функционируют в основном как вставочные нейроны, которые проводят нервные сигналы только на небольшие расстояния в пределах самой коры. Некоторые нейроны — возбуждающие, они выделяют в основном нейромедиатор глутамат; другие — тормозящие, основным непромедиатором которых является гамма-аминомасляная кислота — главный тормозящий нейромедиатор.
Сенсорные области коры, а также ассоциативные, расположенные между сенсорными и моторными областями, содержат большое количество зернистых клеток, что дает основание предполагать высокую степень внутрикоргикалыюй обработки входящих сенсорных сигналов в пределах сенсорных и ассоциативных областеи.
Пирамидные и веретенообразные клетки дают начало почти всем волокнам, выходящим из коры. Пирамидные клетки — более крупные и мно гочисленпые, чем веретенообразные. Они являются источником длинных толстых нервных волокон, которые идут, не прерываясь, в спинной мозг. Они также дают начало крупным подкорковым пучкам ассоциативных волокон, проходящим от одной крупной части головного мозга к другой.
Справа на рисунке выше показана типичная организация нервных волокон в пределах разных слоев коры большого мозга. Обратите особое внимание на большое число горизонтальных волокон, которые распространяются между при лежащими областями коры. Также отметьте вертикальные волокна, идущие к коре и от нее к нижележащим областям мозга. Некоторые из них проходят весь путь до спинного мозга или направляются к отдаленным регионам коры большого мозга через длинные ассоциативные пучки.
Функции специфических слоев коры большого мозга обсуждаются в отдельных статьях на сайте (просим вас пользоваться формой поиска выше). Здесь следует вспомнить, что большинство входящих специфических сенсорных сигналов от тела поступают в IV слой коры. Большая часть сигналов покидают кору через нейроны, локализованные в V и в VI слоях; очень крупные волокна, идущие к мозговому стволу и спинному мозгу, берут начало главным образом в V слое, а громадное количество волокон, идущих к таламусу, возникают в VI слое.
Слои I, II и III осуществляют основную часть ассоциативных функций внутри коры, причем особенно большое число нейронов II и III слоев формируют короткие горизонтальные связи с прилежащими областями коры.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Анатомия коры головного мозга
Толщина коры больших полушарий (от греч. pallium — мантия) варьирует от 2 до 4 мм, наиболее тонкая ее часть находится в области первичной зрительной коры, а наиболее утолщенная — в первичной моторной области. Больше половины общей поверхности коры скрыто стенками борозд. Мозг содержит приблизительно 86 млрд, нейронов (кора больших полушарий содержит только 19% общего количества, но составляет 81% массы мозга), аналогичное число глиальных клеток и разветвленную капиллярную сеть.
Микроскопически кора имеет полосчатый, или пластинчатый, вид, отражающий распределение ее клеток и нервных волокон, а также веерообразную организацию клеточных элементов. Общая цитоархитектоника (схема строения, основанная на типе клеток; схему строения в зависимости от распределения миелинизированных волокон обозначают термином миелоархитектоника) во многом варьирует от зоны к зоне, что позволяет составить «карту» коры с десятками гистологически различных зон. Больших успехов удалось достичь в соотнесении этих зон со специфическими функциями. Несмотря на то, что это имеет теоретическую ценность, подобное разделение довольно упрощено, так как часто принимают во внимание только ключевые функции более обширных функциональных систем, взаимодействующих с другими отделами мозга.
а) Пластинчатое строение. Пластинчатое распределение нейронов характерно для всех отделов коры. Филогенетически «старые отделы», включая древнюю кору (обонятельную кору) и старую кору (гиппокампальная формация и зубчатое ядро; участвуют в процессах памяти) образованы тремя клеточными пластинками. Однако в новой коре (неокортексе или изокортексе, название которой указывает на единый нейрогенез коры, закончившийся образованием шестислойного строения), составляющей оставшиеся 90% (подавляющую часть коры больших полушарий), происходит переход к шестислойному строению.
 Шестислойное строение изокортекса коры головного мозга.
Шестислойное строение изокортекса коры головного мозга.
(А) Соматосенсорная кора. (Б) Первичная моторная кора, кортикальные пластинки обозначены цифрами от I до VI.
Клеточные пластинки новой коры (неокортекса):
I. Молекулярный слой образован концами апикальных дендритов пирамидных клеток и наиболее дистальными ветвями аксонов, проходящих к коре от интраламинарных ядер таламуса.
II. Наружный зернистый слой образован мелкими пирамидными и звездчатыми клетками.
III. Наружный пирамидный слой состоит из средних пирамидных клеток и звездчатых клеток.
IV. Внутренний зернистый слой образован звездчатыми клетками, к которым подходят восходящие волокна от релейных таламических ядер. Звездчатые (зернистые) клетки особенно многочисленны в первичной соматосенсорной коре, первичной зрительной коре и первичной слуховой коре; к ним подходят афферентные чувствительные волокна. Эти отделы обозначают как гранулярная кора. Наоборот, первичная моторная кора — место начала корково-спинномозгового и корково-бульбарного путей, имеет относительно небольшое количество зернистых клеток в слое IV и большое число пирамидных клеток в слоях III и V, которые смазывают (перекрывают) отдельные слои. Эту область обозначают как агранулярная кора.
V. Внутренний пирамидный слой образован крупными пирамидными клетками, волокна которых направляются к полосатому телу, стволу мозга и спинному мозгу.
VI. Слой веретеновидных клеток образован видоизмененными пирамидными клетками, волокна которых направляются к таламусу.
б) Колончатое строение. Помимо хорошо различимой пластинчатой организации коры больших полушарий, можно наблюдать также лучистую, или «колончатую», организацию клеточных структур. Колончатое строение новой коры стало основой исследования функционирования групп нейронов в соматосенсорной коре животных. Подобные лучевидно расположенные группы клеток, вероятно, представляют собой отдельные зоны с одинаковыми физиологическими свойствами и образуют структурную основу для создания более сложных функций. Группы колонн могут образовывать модули, участвующие в обработке различных сенсорных модальностей и функций.
Установлено, что колонны в различных отделах коры имеют разное строение, обусловленное вариабельностью многочисленных свойств, включая строение и число клеток, онтогенез, синаптические связи и молекулярные маркеры. Все это влияет на различные функциональные характеристики и реакции в ответ на возбуждение. В качестве организующего принципа подобная концепция колончатого строения представляет определенную пользу, однако удобнее рассматривать кору как совокупность горизонтального (пластинчатого) и вертикального (лучистого) типов строения. Несмотря на то, что колонны не представляют собой отдельные подобные образования с четкими границами, такая концепция более точно отражает строение коры, а также наблюдаемые экспериментально функциональность, «экономию пространства» и пластичность коры.
Взаимодействие между группами колонн позволяет осуществлять более сложную деятельность, модели поведения или решать когнитивные задачи.
Лежащая в основе организации коры «схема» приводит к тому, что каждая из колонн становится модально (функционально) специфичной после «обработки» информации отдельными элементами. Однако конечная реакция проекционных нейронов колонн может значительно варьировать в зависимости от степени их возбуждения и информации, подходящей к каждому нейрону. Например, одна колонна может реагировать на движение в определенном суставе, но не на раздражение кожи над ним; однако при изменении условий их реакция также может изменяться.
 Восходящие/нисходящие проводящие пути. Стрелки указывают направление проведения импульса. Знаки +/- обозначают возбуждение/торможение.
Восходящие/нисходящие проводящие пути. Стрелки указывают направление проведения импульса. Знаки +/- обозначают возбуждение/торможение.
Пирамидная клетка 1 возбуждается шиповатой зернистой клеткой; она возбуждает клетку 2 внутри своей колонны клеток; клетка 3 в соседней колонне тормозится гладкой зернистой клеткой.
в) Типы клеток. Морфологически кортикальные нейроны разделяют на две большие группы. Большая часть (60-85%) представлена пирамидными нейронами (из-за их формы) — единственными, волокна которых покидают кору (и к которым подходит большая часть волокон), что объясняет их альтернативное название — корковые проекционные нейроны; их волокна — возбуждающие глутаматергические. К оставшимся 15-40% нейронам относят непирамидные, или вставочные, нейроны; несмотря на то, их взаимосвязи не выходят за пределы коры, они осуществляют регуляцию и значительное влияние на ее деятельность; их тип — преимущественно тормозной ГАМК-ергический.
Внутри каждой группы можно выделить множество подгрупп в зависимости от морфологии, взаимосвязей, электрофизиологических свойств, типа развития, физиологических характеристик, молекулярных маркеров и т. д. (Примерами основных морфологических и функциональных клеточных типов служат пирамидные клетки, шиповатые зернистые клетки (измененные пирамидные клетки) и группа непирамидных тормозных вставочных нейронов.)
• Пирамидные клетки имеют пирамидоподобную форму с вершиной, направленной к поверхности. Размеры клеток в высоту составляют 20-30 нм в слоях II и III и более чем в два раза больше в слое V. Самые крупные — гигантские пирамидные клетки Беца — расположены в моторной коре. Единственный апикальный дендрит каждой пирамидной клетки достигает слоя I, заканчиваясь на пучке дендритов. Несколько базальных дендритных ветвей отходят от базальных «углов» клетки и веерообразно расходятся к соответствующим слоям. Апикальные и базальные дендритные ветви свободно разветвляются и усыпаны дендритными шипиками. Большинство пирамидных клеток расположено в слоях II-III и V-VI. Отходящие от основания клеток аксоны отдают несколько возвратных ветвей до вхождения в подлежащее белое вещество, их функция — возбуждение соседних пирамидных клеток.
• Шиповатые зернистые клетки — один из вариантов атипичных пирамидных клеток, лежащих в слое IV и наиболее многочисленных в первичной сенсорной коре. Их шиповатые дендриты не выходят за пределы слоя IV, а аксоны могут подниматься или опускаться, образуя возбуждающие глутаматергические синаптические контакты с пирамидными клетками. На них переключается большая часть волокон восходящих таламических путей к слою IV с дальнейшим радиальным распространением.
Нейромедиатором различных по структуре и классификации непирамидных тормозных вставочных нейронов служит ГАМК. [Сложная классификация нейронов новой коры постоянно меняется. Гладкие звездчатые (зернистые) клетки можно обнаружить во всех слоях коры; их дендриты веерообразно расходятся во всех направлениях, а их аксоны образуют локальные древовидные разветвления, поэтому их иногда называют нейронами локальных сплетений. Несмотря на их уникальные морфологические характеристики, нейроглиеформные, клетки-канделябры и корзинчатые клетки считают специализированными типами зернистых клеток. Наш совет: если Вы встречаете термины «зернистая» или «гладкая звездчатая» клетка, для облегчения чтения и понимания их следует рассматривать как вставочные нейроны.]
Для организации их можно разделить на три крупных семейства в зависимости от экспрессируемых этими вставочными нейронами биомаркеров: парвальбумин, соматостатин и серотонин (5-гидрокситриптамин, 5НТ) 3а-рецептор (5HT3aR).
• Парвальбумин-экспрессирующие вставочные нейроны не имеют шигшков на дендритах. К ним подходят возбуждающие волокна от таламуса и коры, а тормозные — от других вставочных нейронов того же типа. Считают, что они играют роль в стабилизации активности корковых нейронных сетей. Как и в коре мозжечка, эти нейроны обладают фокусирующим действием на кору больших полушарий, тормозя слабо возбужденные колонны клеток. Клетки-канделябры (названы так из-за канделяброподобных групп аксоаксональных синаптических окончаний) наиболее многочисленны в слое II, образуют контакты с начальным сегментом аксона пирамидной клетки и играют ключевую роль в корково-корковых взаимодействиях.
Корзинчатые клетки лежат преимущественно в слоях II и V, а свое название получили благодаря тому, что их аксоны формируют околоклеточные «корзинки» вокруг тел пирамидных клеток, дистальных отделов их дендритов и аксонов других корзинчатых клеток.
• Соматостатин-экспрессирующие вставочные нейроны. Примером служат клетки Мартинотти, расположенные в пластинках V и VI, аксоны которых направляются к пластинке I. Получая сигналы от пирамидных клеток, они способны ограничивать их периферическое возбуждение и объединять несенсорную информацию, осуществляя, таким образом, регулирование обработки сигналов в дендритах их пирамидных клеток в зависимости от внешних обстоятельств.
• 5НТ3а-экспрессирующие вставочные нейроны — гетерогенная группа, к которой относят большую часть вставочных нейронов поверхностных корковых слоев. Получая информацию от корковых и таламических нейронов, они могут участвовать в процессах обучения, осуществляя влияние на корковые нейронные цепи. Он нейроглиеформных клеток (паутинообразных клеток)—одного из главных типов вставочных нейронов слоев II и III — веерообразно расходятся дендриты, обладающие уникальным свойством образования синапсов друг с другом и другими типами вставочных нейронов; этот факт указывает на их важнейшую роль в синхронизации корковых нейронных цепей. Другая морфологически гетерогенная группа вставочных нейронов, помимо ГАМК, экспрессирует вазоактивный интестинальный полипептид; другие типы вставочных нейронов этой группы экспрессируют также холецистокинин и другие пептидные рецепторы.
 Три морфологических типа ГАМК-ергических тормозных нейронов:
Три морфологических типа ГАМК-ергических тормозных нейронов:
А. Аксодендритическая клетка, образующая синапс с апикальным отделом пирамидного нейрона.
Б. Корзинчатая клетка, образующая аксоаксональные синапсы с пирамидными клетками.
В. Клетка-канделябр, формирующая аксоаксональные синапсы (*) с начальными сегментами аксонов двух пирамидных клеток, показанных здесь, и с начальными сегментами четырех других клеток, не показанных на рисунке.
(На основе DeFelipe, 1999, с разрешения автора.)
г) Афферентные волокна. Восходящие волокна к любому отделу коры могут иметь четыре различных источника (в первую очередь, корковые) и оканчиваются в разных отделах.
1. Длинные и короткие ассоциативные волокна от мелких и средних пирамидных клеток в слоях II и III из других корковых зон того же полушария.
2. Комиссуральные волокна от средних пирамидных клеток слоев II и III проходят через мозолистое тело от аналогичных или топографически идентичных участков противоположного полушария.
3. Таламо-корковые волокна от соответствующего специфического или ассоциативного ядра (например, волокна от вентрального заднего таламического ядра к соматосенсорной коре) и от дорсомедиального таламического ядра к предлобной (префронтальной) коре (описана ниже) оканчиваются в пластинке IV. Неспецифические таламо-корковые волокна от внутрипластинчатых ядер оканчиваются во всех пластинках.
4. Холинернические и аминергические волокна от базальных ядер, гипоталамуса и ствола мозга. Эти волокна обозначены на рисунке ниже зеленым цветом. Несмотря на то, что они распространены по всей коре, их возбуждение не сопровождается генерализованной или неспецифической реакцией. Анатомическая специфичность волокон (корковая, пластинчатая и клеточная) обусловливает возбуждение или торможение только определенных групп нейронов. Ядра их начала и нейромедиаторы:
• базальное ядро Мейнерта (базальные ядра мозга), ацетилхолин;
• серобугорно-сосцевидное ядро (задний отдел гипоталамуса), гистамин;
• черная субстанция, компактный слой (вентральный отдел покрышки среднего мозга), дофамин;
• ядра шва (средний мозг и ростральный отдел моста), серотонин;
• голубое пятно (ростральный отдел моста), норадреналин. Эти пять групп нейронов имеют особое значение в психиатрии и подробно рассмотрены в отдельной статье на сайте.
д) Эфферентные волокна. Единственные эфферентные волокна, покидающие кору больших полушарий, представлены возбуждающими аксонами пирамидных клеток. Одна часть аксонов пирамидных клеток соединяется с длинными и короткими ассоциативными волокнами, другая образует комиссуральные или проекционные пути. Ассоциативные и комиссуральные пути составляют большой объем белого вещества полушарий головного мозга.
• Примерами коротких ассоциативных путей (проходят между соседними зонами коры в составе поверхностного белого вещества в виде U-волокон) служат пути, направляющиеся в моторную кору от сенсорной коры и обратно. Примерами длинных ассоциативных путей служат волокна между префронтальной корой (лежащей кпереди от моторных зон коры) и ассоциативными сенсорными зонами. Источник этих волокон — пирамидные клетки, расположенные преимущественно в слоях II и III.
• Комиссуральные волокна мозга представлены исключительно аксонами пирамидных клеток, проходящих через мозолистое тело, переднюю и заднюю спайки (и другие мелкие спайки) к аналогичным участкам противоположного полушария (например, волокна от первичной корковой зоны направляются к соответствующей ассоциативной области другого полушария), а также к несоответствующим зонам [подобные комиссуральные связи отсутствуют между первичной зрительной корой, первичной соматосенсорной и моторной корой, иннервирующими дистальный отдел верхней конечности]. Волокна начинаются от пирамидных клеток, расположенных преимущественно в слоях II и III.
• Основная часть проекционных волокон от первичной сенсорной и моторной коры подходит к базальным ганглиям. К таламусу направляются волокна от всех отделов коры. К другим основным проводящим путям относят корково-мостовой (к ипсилатеральным ядрам моста), корково-ядерный (к двигательным и чувствительным ядрам черепных нервов моста и продолговатого мозга на противоположной стороне) и корково-спинномозговой. Источником этих волокон служат пирамидные клетки, расположенные преимущественно в слоях V и VI (отдающие волокна преимущественно к специфическим релейным ядрам таламуса).
Редактор: Искандер Милевски. Дата публикации: 22.11.2018