в коре головного мозга различают зоны
В коре головного мозга различают зоны
Наиболее широко используют карту полей Бродмана, на которой кора разделена на 44 цитоархитектонические зоны (его схема была расширена до 52 полей, однако не все поля использовали). Большая часть полей представлена на рисунке ниже, однако четких границ между этими полями не существует. (Эти числа часто используют для обозначения функциональных областей, хотя Бродман отвергал подобную корреляцию.)
На рисунке ниже разным цветом выделены три основные первичные сенсорные зоны (соматическая, зрительная и слуховая) и одна первичная моторная зона. Рядом с каждой первичной сенсорной или моторной зоной расположена ассоциативная кора—унимодальная ассоциативная зона (той же модальности). Остальная часть новой коры представлена мультимодальными (полимодальными) ассоциативными зонами, к которым подходят волокна от многих унимодальных ассоциативных зон (например, от зон тактильной и зрительной чувствительности, зрительной и слуховой) и других мультимодальных или паралимбических областей.
 Цитоархитектонические поля Бродмана. Окрашенные зоны:
Цитоархитектонические поля Бродмана. Окрашенные зоны:
а) Моторная (красная):
4 — первичная моторная кора
6 — на медиальной поверхности, дополнительная моторная зона
6 — на латеральной поверхности, премоторная кора
б) Сенсорная (голубая):
3/1/2 — первичная соматосенсорная кора
40 — вторичная соматосенсорная кора
17 — первичная зрительная кора 18,
19 — ассоциативная зрительная кора
41, 42 — первичная слуховая кора*
22 — ассоциативная слуховая кора
(*Первичную слуховую кору не всегда можно увидеть сбоку, так как она полностью расположена на верхней поверхности верхней височной извилины.)
Изучение функциональной анатомии. Термином коннектом обозначают «полную карту нейронных связей, охватывающих все функции мозга». Однако для создания законченной функциональной карты человеческого мозга требуется объединить эмпирические данные со структурными связями, при этом многое все еще остается неизвестным. Современные подходы создают уникальные возможности для достижения этой цели с помощью новых возможностей обработки и хранения данных, нейрофизиологических исследований и магнитно-резонансной томографии (МРТ), позволяющих получить изображения головного мозга живого человека.
Новые достижения в понимании устройства мозга ознаменовались смещением приоритета с отдельных зон коры на рассмотрение всех отделов и взаимосвязей как единого целого. Были разработаны новые теоретические и методологические схемы, позволяющие описывать и прогнозировать сложную системную динамику путем использования сетевого анализа и математических методов, основанных на теории графов. В сетевых моделях используют совокупности «элементарных» корковых единиц и их взаимодействия, чтобы показать появление функционирующих участков в динамике или «поймать мозг в действии».
Эти модели остаются ограниченными известными взаимосвязями между зонами коры, а существование некоторых взаимосвязей было предположено по результатам исследований у приматов. Однако данные модели позволяют предположить наличие взаимосвязей или проводящих путей, существование которых структурно не доказано, на основании типа реакции. Несмотря на прогресс в исследовании проводящих путей и взаимосвязей в коре при лучевой диагностике мозга живого человека благодаря использованию нейрорентгенологических методов и математического моделирования, внедрение новых и продолжение использования «старых» техник изучения нейроанатомии необходимо для получения структурных свидетельств существования этих проводящих путей и возбуждающих нейронных систем.
Для «определения и локализации» функций головного мозга используют преимущественно две методики. В основе обеих лежит регистрация локального усиления кровотока в ответ на повышенную потребность мозга в кислороде в результате увеличения нейронной активности.
1. Позитронная эмиссионная томография (ПЭТ). С помощью позитронной эмиссионной томографии (ПЭТ) измеряют потребление кислорода после введения в вену предплечья воды, меченной кислородом-15 ( 15 O). 15 O — излучающий позитроны изотоп кислорода; в крови позитроны реагируют с окружающими электронами и испускают у-лучи, которые регистрируют детекторы γ-лучей. Для измерения уровня потребления глюкозы используют также 18-фтордезоксиглюкозу ( 18 F-дезок-сиглюкоза). Нейроны захватывают 18 F-дезоксиглюкозу в той же степени, что и глюкозу.
Для правильной интерпретации данных ПЭТ требуется выполнение вычитания изображений и усреднения изображений, описанных под рисунком ниже. Аналогичная методика извлечения сигнала описана для функциональной МРТ (фМРТ).
При проведении специализированных исследований (например, при определении суммарной функции рецепторов) используют меченные изотопом химические вещества: меченный радиоизотопом дофамин в полосатом теле при болезни Паркинсона, меченный радиоизотопом серотонин в стволе мозга и коре больших полушарий при депрессии, меченую ацетилхолинэстеразу при болезни Альцгеймера.
2. Функциональная магнитно-резонансная томография (фМРТ). При фМРТ не требуется введения дополнительных веществ. Метод основан на различии в магнитных свойствах оксигенированной и неоксигенированной крови. Если локального усиления кровотока более чем достаточно для покрытия потребности в кислороде, повышается коэффициент отношения оксигемоглобина к дезоксигемоглобину, что ведет к образованию MPT-сигнала. Функциональные и структурные взаимосвязи можно выявить при взаимном изменении или колебании интенсивности фМРТ-сигнала в различных корковых зонах даже при отсутствии «прямых» кортикальных связей. Приведенные ниже данные были получены с помощью функциональных методов исследования, клинических наблюдений и результатов экспериментов на животных.
 Вычитание изображений и усреднение изображений при позитронной эмиссионной томографии (ПЭТ).
Вычитание изображений и усреднение изображений при позитронной эмиссионной томографии (ПЭТ).
Верхний. Контрольное среднее изображение получено у пациента в покое. Захват 15 O происходит во всех отделах коры и подкоркового серого вещества.
Левое изображение получено у того же пациента, следящего за движущимися по экрану точками.
Высокий уровень фоновой активности скрывает результат исследования. Правое изображение получено вычитанием контрольного изображения и позволяет увидеть повышенную активность зрительной коры при выполнении зрительной задачи.
Средний. Аналогичную задачу выполняли другие четыре пациента. Вычитание фонового «шума» позволило выявить значительные различия между пациентами.
Поскольку размеры мозга у людей варьируют, активность мозга у этих пятерых пациентов была наложена на общий, «средний» мозг (хотя мозг на всех рисунках изображен одинаково). Нижний. Усредненное значение пяти исследований указывает на среднюю разницу в этой группе.
Редактор: Искандер Милевски. Дата публикации: 22.11.2018
Кора головного мозга: участки, анализаторы
Значение, роль коры больших полушарий головного мозга человека
В статье мы рассмотрим локализацию функций, участки, анализаторы, поля, участки, области зоны коры больших полушарий головного мозга человека (мужчины, женщины). Неврологи, невропатологи, рефлексотерапевты, рефлексологи выделяют 4 основных положения, применительно к практической деятельности невропатолога, современного учения о локализации функций в коре головного мозга.
1. Очень сложная морфологическая и функциональная дифференциация коры больших полушарий головного мозга. Лобная доля больше отвечает за двигательные функции. Теменная, затылочная и височная зоны больше отвечают за чувствительные функции.
3. Формирование специальных корковых областей в процессе практической деятельности.
Функция творит центр
По Ивану Петровичу Павлову: «Функция творит центр!» В раннем детстве границы корковых центров диффузны и менее дифференцированы, и лишь по мере приобретения жизненного опыта происходит постепенная концентрация функциональных зон, в связи с чем у детей первых лет жизни слабо выражены очаговые корковые симптомы и чаще преобладает общемозговая симптоматика.
4. Существенные различия в локализации более простых и более сложных функций. Чем проще функция, тем она точнее локализована. И наоборот, наиболее сложные функции обусловлены интегративной деятельностью всего головного мозга, поэтому понятие «корковый центр» (отдел коры головного мозга, поля коры головного мозга, участки коры головного мозга, части коры головного мозга) в большинстве случаев относительное и условное. К простым корковым функциям относятся чувствительная функция, двигательная функция, зрительная функция, слуховая функция, вестибулярная функция, обонятельная функция, вкусовая функция. К сложным корковым функциям относятся речь, письмо, чтение, счет, праксис, гнозис, мышление, память.
Локализация функций и симптомов
Проводя топическую диагностику рефлексотерапевт, невролог, невропатолог, микроневропатолог, детский невролог, взрослый невролог определяет не только локализацию поражения корковых центров, но и локализацию симптомов. Простые корковые функции связаны с проекционными пластинками коры (пятой и четвертой), имеющими непосредственную связь с периферией и являющимися корковыми отделами анализаторов. Сложные корковые функции связаны с ассоциативными слоями коры (вторым и третьим). Последние слои соединены горизонтальными волокнами с другими участками коры головного мозга в пределах одного полушария и не имеют прямого выхода на периферию. Большое значение в обеспечении сложных корковых функций имеют также комиссуральные связи между полушариями, проходящими через мозолистое тело.
Простые корковые функции обычно представлены в обоих полушариях головного мозга. Сложные корковые функции чаще имеют асимметричное представительство в правом или левом полушарии головного мозга. Итак, какие бывают поля, участки, области, типы коры головного мозга, отделы, анализаторы, части коры головного мозга?
Двигательная кора головного мозга, двигательные центры головного мозга, двигательные анализатор, моторный
Главным корковым отделом двигательного анализатора, его первичным полем, является предцентральная извилина, в верхних отделах которой находится проекционная область мышц стопы, голени, бедра, в средней части – туловища и руки, в нижней трети – лица. Двигательная иннервация построена по соматотопическому принципу. На этом уровне осуществляются тонкие дифференцированные движения. Кроме того, имеются дополнительные двигательные зоны – это вторичные поля двигательного анализатора и третичные поля двигательного анализатора. Дополнительные двигательные зоны обеспечивают сложные автоматизированные двигательные акты. Например, в парацентральной дольке находятся корковые центры тазовых органов. В задних отделах верхней лобной извилины находится переднее адверсивное поле. Заднее адверсивное поле располагается на границе верхней теменной дольки и затылочной области. Задние отделы средней лобной извилины отвечают за сочетанный поворот головы и глаз в противоположную сторону. Задние отделы нижней лобной извилины осуществляет движения типа орального автоматизма – глотание, жевание, лизание.
Чувствительная кора головного мозга, чувствительные центры головного мозга, чувствительный анализатор
Главным корковым отделом поверхностных и глубоких видов чувствительности является постцентральная извилина, где также имеется соматотопическое представительство участков периферии, аналогичное вышеуказанному. К поверхностной чувствительности относятся температурная чувствительность, болевая чувствительность, тактильная чувствительность.
Стереогноз, стереогнозис
Сложные виды чувствительности локализованы в коре полушарий головного мозга на уровне верхней теменной дольки, где отсутствует соматотопика. К сложным видам чувствительности относятся стереогностическая чувствительность (стереогноз, стереогнозис), двумерно-пространственная чувствительность, чувство локализации и дискриминации. Зрительная проекционная зона (зрительная зона коры) занимает область шпорной борозды – внутренняя поверхность затылочной доли. Слуховая проекционная зона (слуховая зона коры) занимает центр верхней височной извилины и извилину Гешля. Вестибулярная проекционная зона находится рядом со слуховой. Обонятельная проекционная зона локализуется на внутренней поверхности височной доли, в извилине гиппокампа. Вкусовая проекционная зона находится рядом с последней, а также в области покрышки и островка Reili.
Теперь остановимся на локализации сложных корковых функций.
Обычно сложные корковые функции локализуются в левом полушарии головного мозга у правшей и в правом полушарии головного мозга у левшей.
Функцию речи обеспечивает сенсорный центр (центр Вернике), который располагается в заднем отделе верхней височной извилины. При поражении центра Вернике наблюдается сенсорная афазия. Также функцию речи обеспечивает двигательный центр (центр Брока), который располагается в области задних отделов нижней лобной извилины. При поражении центра Брока наблюдается моторная афазия. При патологии на стыке височной и затылочной долей формируется амнестическая афазия и семантическая афазия. Речевые зоны коры головного мозга.
Лексический анализатор, центр лексии, функция чтения
Функции чтения обеспечивает лексический центр (центр лексии). Центр лексии располагается в угловой извилине.
Графический анализатор, центр графии, функция письма
Функции письма обеспечивает графический центр (центр графии). Центр графии располагается в заднем отделе средней лобной извилины.
Счетный анализатор, центр калькуляции, функция счета
Функции счета обеспечивает счетный центр (центр калькуляции). Центр калькуляции располагается на стыке теменно-затылочной области.
Праксис, праксический анализатор, центр праксиса
Праксис – это способность к выполнению целенаправленных двигательных актов. Праксис формируется в процессе жизнедеятельности человека, начиная с грудного возраста, и обеспечивается сложной функциональной системой мозга с участием корковых полей теменной доли (нижняя теменная долька) и лобной доли, особенно левого полушария у правшей. Для нормального праксиса необходимы сохранность кинестетической и кинетической основы движений, зрительно-пространственной ориентировки, процессов программирования и контроля целенаправленных действий. Поражение праксической системы на том или ином уровне проявляется таким видом патологии, как апраксия. Термин «праксис» происходит от греческого слова «praxis», которое означает «действие». Апраксия – это нарушение целенаправленного действия при отсутствии параличей мышц и сохранности составляющих его элементарных движений.
Гностический центр, центр гнозиса
В правом полушарии у правшей, в левом полушарии головного мозга у левшей представлены многие гностические функции. При поражении преимущественно правой теменной доли может возникать анозогнозия, аутопагнозия, конструктивная апраксия. С центром гнозиса также связаны музыкальный слух, ориентация в пространстве, центр смеха.
Память, мышление
Наиболее сложные корковые функции – это память и мышление. Эти функции не имеют четкой локализации.
Память, функция памяти
Мышление, функция мышления
Функция мышления – это результат интегративной деятельности всего головного мозга, особенно лобных долей, которые участвуют в организации целенаправленной сознательной деятельности человека, мужчины, женщины. Происходят программирование, регуляция и контроль. При этом у правшей левое полушарие является основой преимущественно абстрактного словесного мышления, а правое полушарие связано главным образом с конкретным образным мышлением.
Развитие корковых функций начинается с первых месяцев жизни ребенка, достигает своего совершенства к 20 годам.
Зоны коры головного мозга
В последующих статьях мы остановимся на актуальных вопросах неврологии: зоны коры головного мозга, зоны больших полушарий, зрительная, зона коры, слуховая зона коры, моторные двигательные и чувствительные сенсорные зоны, ассоциативные, проекционные зоны, моторные и функциональные зоны, речевые зоны, первичные зоны коры головного мозга, ассоциативные, функциональные зоны, фронтальная кора, соматосенсорная зона, опухоль коры, отсутствие коры, локализация высших психических функций, проблема локализации, мозговая локализация, концепция динамической локализации функций, методы исследования, диагностики.
В коре головного мозга различают зоны
а) Первичная моторная кора. Первичная моторная кора (поле 4) — полоска агранулярной коры в прецентральной извилине. Она дает начало 60-80% (количество варьирует) волокон корково-спинномозгового пути (КСП). Остальная часть волокон отходит от премоторной, поясной и добавочной моторной зон, а также теменной коры, что показано в отдельной статье на сайте. Наибольшую плотность окончаний КСП в спинном мозге наблюдают в зонах, иннервирующих дистальные мышцы конечностей.
Части тела противоположной стороны соматотопически представлены в коре инвертированными, за исключением лица, а относительно крупные области занимают кисть (играет важную роль в контроле движений пальцев), околоротовая область и язык. Зону кисти обычно можно определить по направленному кзади возвышению в 6-7 см от верхнего края полушария.
Ипсилатералъные части тела также имеют соматотопическую организацию; ипсилатеральные моторные нейроны составляют лишь 10% волокон пирамидного пути, которые не переходят на противоположную сторону и не иннервируют дистальные мышцы конечностей.
При целенаправленной стимуляции моторной коры человека установлено, что клеточные колонны регулируют направление движения. Первичная моторная кора «синтезирует» двигательные команды, но не является местом их возникновения. От нее волокна в составе КСП направляются в спинной мозг и широко разветвляются по мере достижения места своего окончания. Для того чтобы взять, например, ручку в руку, требуется:
(а) умеренное сокращение мышцы, противопоставляющей большой палец, как основной движущей мышцы;
(б) сокращение с той же интенсивностью отдела глубокого сгибателя пальцев, от которого отходит сухожилие к дистальной фаланге указательного пальца;
(в) более слабое сокращение мышцы, приводящей большой палец, и
(г) короткого сгибателя большого пальца.
Удержание всей руки при любом типе манипуляций — функция премоторной коры, отражающая важность бессознательной регуляции положения тела при совершении произвольных движений. Более крупные моторные зоны коры формируются при «объединении» близлежащих нейронных колонн на основе их функций и создании сложных последовательностей движений.
1. Пластичность моторной коры. Некрупные поражения моторной коры у обезьян и низших млекопитающих приводят первоначально к параличу соответствующей части тела, а затем в течение нескольких дней (иногда часов) — к быстро прогрессирующему восстановлению функции. Восстановление может быть обусловлено изменением свойств клеточных колонн, расположенных рядом с местом поражения, которые берут на себя утраченную двигательную функцию. Увеличение двигательной зоны участка коры возможно путем локального введения антагонистов ГАМК в кору. Расширение моторной зоны на уровне спинного мозга заранее обеспечено обширными зонами перекрытия от поля 4 до колонок двигательных нейронов серого вещества переднего рога, однако степень пластичности здесь ниже, чем в коре.
Взаимосвязи между КСП (и другими нисходящими путями) и двигательными нейронами в спинном мозге возникают посредством вставочных нейронов. Во вставочных нейронах происходит объединение сенсорной и корковой информации, что сопровождается специфической и организованной активацией пулов двигательных нейронов и сокращением иннервируемых ими мышц.
2. Источники афферентных путей к первичной моторной коре:
— Моторная кора противоположной стороны через мозолистое тело. Наиболее прочные комиссуральные связи возникают между соответствующими колоннами клеток, иннервирующими мышцы живота и спины. Это вполне понятно, поскольку данные группы мышц обычно сокращаются одновременно с двух сторон, поддерживая вертикальное положение головы и туловища. Самые слабые комиссуральные связи имеются между колоннами клеток, контролирующими дистальные группы мышц конечностей, движения в которых слева и справа происходят независимо друг от друга.
— Соматосенсорная кора. Кожные колонны клеток в полях 1, 2 и 3 отдают кпереди короткие ассоциативные волокна (взаимосвязи с зоной кисти особенно многочисленные; эти волокна короткие, так как зоны, отвечающие за кисть в сенсорной и моторной коре, обычно занимают соседние стенки центральной борозды). Проприоцептивные клеточные колонны получают афферентые волокна от аннуло-спиральных окончаний мышечных веретен; они отдают короткие ассоциативные волокна к соответствующим двигательным колоннам, активируя рефлексы растяжения с длинной дугой.
— Противоположное зубчатое ядро. Мозжечок принимает участие в выборе нужных мышц-синергистов, а также времени и силы их сокращений.
— Дополнительная моторная зона (ДМЗ).
 (А) Предполагаемая функциональная организация сегмента запястье-кисть в первичной двигательной коре (М1) у обезьян и других приматов.
(А) Предполагаемая функциональная организация сегмента запястье-кисть в первичной двигательной коре (М1) у обезьян и других приматов.
Несмотря на то, что М1 имеет в целом соматотопическую организацию, локальная соматотопическая структура разбита в виде мозаики на радиальные ряды нейронов, инициирующих небольшие специфические движения.
Миниколонны, отвечающие за движения отдельных пальцев, могут примыкать к колоннам запястья, локтя или плеча, а подгруппы этих миниколонн объединены по функции образования более сложных последовательностей движений, таких как хватание, вытягивание руки или защита рукой от удара.
(Б) Пространственная организация моторных нейронов коры приматов, контролирующих движения мышечных групп.
От медиальной стороны к латеральной они располагаются в следующей последовательности: стопа, голень, плечо, кисть и голова (голубой).
В каудальном отделе М1 лежат двигательные нейроны, взаимодействующие непосредственно с двигательными нейронами (спинного мозга) и отвечающие за высокоточные движения, необходимые для выполнения тонких двигательных навыков.
Эти нейроны расположены в медиально-латеральной последовательности от проксимальных (красный) до дистальных (желтый) групп мышц. Авторы обозначают эту зону как «новую М1», подотдел «старой М1» (голубая) для того, чтобы отразить недавнее возникновение сложной двигательной активности и эволюционно «нового» отдела моторной коры.
б) Премоторная кора. Премоторная кора (поле 6 на латеральной поверхности полушария) примерно в 6 раз крупнее первичной моторной коры. Она получает когнитивные импульсы от лобной доли при намерении произвести движение, а также от теменной доли (поле 7), передающей огромное количество тактильной и пространственно-зрительной информации. Максимальная ее активация происходит, когда последовательность движений происходит в ответ на зрительные или соматосенсорные раздражители, такие как приближение к объекту, полностью находящему в поле зрения, или идентификация объекта, находящегося вне поля зрения путем ощупывания. Активация премоторной коры, как правило, двусторонняя. Одним из объяснений служит межполушарная передача двигательных программ через мозолистое тело.
Следует также принимать во внимание обширные связи премоторной коры с ядрами ствола мозга, дающими начало ретикуло-спинномозговым путям (и небольшому пути к КСП). Поражения, затрагивающие премоторную кору у человека, встречают редко, они характеризуются нестабильностью положения в противоположном плече и бедре. Таким образом, важной функцией премоторной коры, вероятно, служит двустороннее поддержание позы, например фиксация плечей при работе двумя руками и стабилизация бедер при ходьбе. Премоторная кора может вносить вклад в восстановление функции при истинной моторной гемиплегии, возникающей вследствие сосудистого поражения, затрагивающего КСП в лучистом венце. При данном поражении во время ПЭТ-исследования наблюдают повышение активности премоторной коры; кпереди от КСП спускается корково-ретикуло-спинномозговой проводящий путь.
в) Дополнительная моторная зона (ДМЗ). В отличие от премоторной коры, реагирующей на внешние раздражители, ДМЗ (поле 6 на медиальной поверхности полушария) отвечает на внутренние раздражители и принимает особое участие в планировании движений. Это подтверждает тот факт, что ДМЗ возбуждается лобной долей (ДЛПФК) в тот момент, когда мы собираемся совершить движение, даже если движение в итоге не возникает. Важнейшей функцией ДМЗ, вероятно, является предварительное программирование последовательностей движений, уже заложенных в двигательной памяти. ДМЗ функционирует совместно с моторной петлей, проходящей через базальные ганглии, а также взаимодействует с полем 4 и отдает волокна непосредственно в КСП.
Одностороннее поражение ДМЗ может сопровождаться акинезией (невозможностью начать движение) руки и ноги с разных сторон. Двустороннее поражение приводит к тотальной акинезии, включая речевую акинезию.
1. Корковые глазодвигательные поля. На рисунке ниже показаны корковые глазодвигательные поля, участвующие в сканирующих движениях (саккадах). Их взаимодействия и функции представлены в таблице ниже.
2. Дорсолатеральная префронтальная кора (ДЛПФК). ДЛПФК — высший когнитивный центр, участвует в оценке зрительной картины, принятии решения о сознательном совершении саккад и добровольном подавлении рефлекторных саккад (произвольные саккады возникают по воле человека). Рефлекторные саккады — автоматические реакции на объекты, появляющиеся в периферическом поле зрения. Строго говоря, рефлекторные саккады следует называть ответными; они не являются истинными рефлексами, так как способны подавляться волей.
3. Поясная кора. Вместе с ДЛПФК участвует в принятии решений и оценке эмоциональной важности, или ценности, видимых объектов.
4. Дополнительное глазодвигательное поле. Занимает передний отдел ДМЗ и участвует в планировании движений, особенно, если требуются множественные саккадические движения.

5. Фронтальное глазодвигательное поле. Фронтальное глазодвигательное поле (ФГДП) инициирует произвольные саккады, благодаря которым происходит переключение внимания на раздражитель или подавляется стремление к прямому взору на новый раздражитель в ответ на одно или более из трех перечисленных афферентных влияний. ФГДП «поддерживает» карту зрительного пространства относительно глазодвигательных координат и вместе с верхним холмиком играет ключевую роль в создании зрительно-направленных и произвольных саккад; поражения обеих структур приводят к необратимой утрате саккадических движений. Клинические и экспериментальные наблюдения у обезьян показали следующее.
• ФГДП имеют тоническую двустороннюю активность.
• Повышение активности среднего отдела ФГДП с одной стороны приводит к образованию горизонтальных саккад, направленных в сторону противоположного зрительного поля (противоположной саккады).
• Повышение активности верхнего отдела с одной стороны приводит к созданию нисходящей косонаправленной саккады; двусторонняя активация верхнего отдела приводит к обращению взора обоих глаз вертикально вниз.
• Повышение активности в нижнем отделе приводит к аналогичным последствиям относительно взора вверх.
6. Теменное глазодвигательное поле. Теменное глазодвигательное поле (ТГДП) отвечает за создание рефлекторных саккад и побуждает ФГДП к созданию произвольных саккад. ТГДП также участвует в пространственном восприятии, создавая схему зрительной картины.
Поражения префронтальной коры и лобной доли описаны в отдельной статье на сайте.
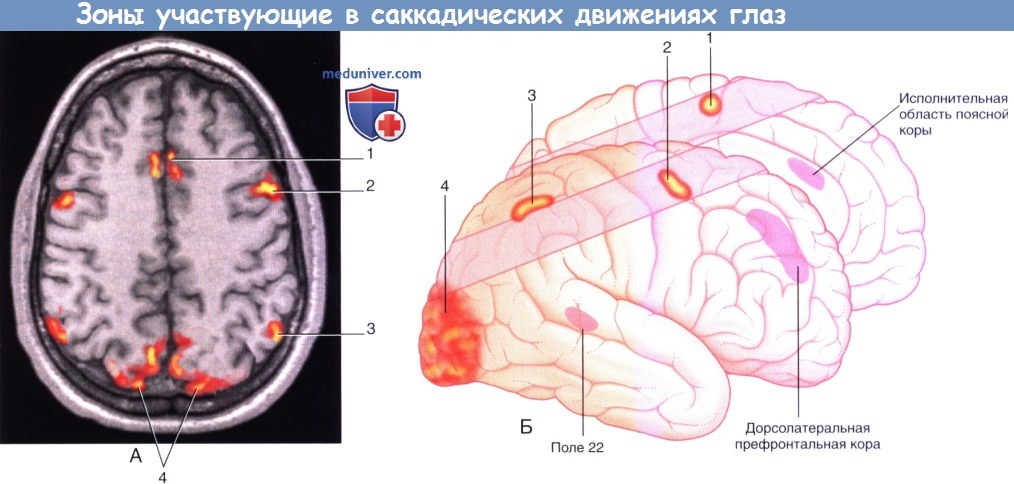 Зоны коры больших полушарий, участвующие в саккадических движениях глаз.
Зоны коры больших полушарий, участвующие в саккадических движениях глаз.
(1) Дополнительное глазодвигательное поле.
(2) Фронтальное глазодвигательное поле.
(3) Теменное глазодвигательное поле.
(4) Ассоциативная зрительная кора.
в) Синдром ригидного человека (СРЧ). Редкое, но легко распознаваемое заболевание, известное как синдром ригидного человека (СРЧ), или синдром мышечной скованности, — аутоиммунное заболевание центральной нервной системы, связанное с наличием циркулирующих антител к глутаматдекарбоксилазе (GAD65),— ключевому ферменту, преобразующему глутамат в ГАМК. Синдром ригидного человека (СРЧ) проявляется скованностью мышц с эпизодическими мышечными спазмами (вызванными одновременным сокращением мышц синергистов и антагонистов, преимущественно в проксимальных отделах конечностей и осевой мускулатуре) и боязнью выполнения определенных задач. В норме контроль верхних моторных нейронов происходит за счет тонической активности прилежащих тормозных ГАМК-ергических вставочных нейронов.
Одни зоны коры поражаются сильнее других, а клинические проявления связаны с нарушением функции этих ГАМК-ергических нейронов, что приводит к гипервозбудимости коры. В настоящее время уточняют роль циркулирующих антител в патогенезе синдрома ригидного человека (СРЧ).
г) Резюме. Кора больших полушарий имеет одновременно пластинчатую и колончатую организацию. К двум основным типам клеток относят пирамидные и непирамидные клетки (вставочные нейроны). Пирамидные клетки расположены в слоях (пластинках) II, III, а также V (в виде веретеновидных клеток) и VI. Слой IV богат шиповатыми звездчатыми клетками (модифицированными пирамидными нейронами). Мелкие пирамидные клетки связывают извилины одного полушария, средние пирамидные клетки связывают между собой соответствующие зоны двух полушарий, а от крупных пирамидных клеток отходят волокна к таламусу, стволу мозга и спинному мозгу. Все корковые волокна считают возбуждающими; шиповатые звездчатые клетки также считают возбуждающими пирамидными клетками.
Корковые вставочные нейроны—тормозные. Колончатая организация представлена колоннами клеток, которые рассматривают в качестве первичной функциональной единицы обработки информации в коре; они состоят из определенных нейронных «микроцепей».
Части тела в соматосенсорной коре представлены инвертированными. Важная входящая информация поступает от вентрального заднего ядра таламуса, а направляется к первичной моторной коре и нижней теменной коре. В первичную зрительную кору входит коленчато-шпорный путь. Клеточные реакции различной сложности зависят от передачи информации с более простых типов клеток на более сложные. Свойство ассоциативных зрительных зон—распознавание черт, например цвета, формы и движения. Обработка цвета и формы продолжается в коре нижележащей височной доли, а движений—в задней теменной доле. Первичная слуховая кора занимает верхнюю поверхность верхней височной извилины, а ассоциативная слуховая кора расположена с латеральной стороны от нее.
Первичная моторная кора занимает прецентральную извилину. Она дает начало большей части волокон пирамидного тракта, а части тела в ней представлены инвертированными. Основная информация подходит к ней от соматосенсорной коры, мозжечка (через вентральное заднее ядро таламуса), премоторной и дополнительной моторной зон. Премоторная кора отвечает преимущественно на внешние раздражители, а дополнительная моторная зона — на внутренние стимулы. Дорсолатеральная префронтальная кора контролирует четыре различных корковых зоны, в различной степени участвующих в создании противоположных саккад.
Редактор: Искандер Милевски. Дата публикации: 22.11.2018