В толще белого вещества каждого полушария большого мозга имеются скопления серого вещества, образующего отдельно лежащие базальные ядра, которые залегают ближе к основанию мозга. К ним относятся полосатое тело (хвостатое и чечевицеобразное ядра), ограда и миндалевидное тело.
Полосатое тело на разрезах мозга имеет вид чередующихся полос серого и белого вещества. Наиболее медиально и впереди находится хвостатое ядро, расположенное латеральнее и выше таламуса, будучи отделенным от него коленом внутренней капсулы.
Хвостатое ядро имеет головку, залегающую в лобной доле, выступающую в передний рог бокового желудочка. Передняя ножка внутренней капсулы (белое вещество) отделяет хвостатое ядро от чечевицеобразного ядра. Тело хвостатого ядра лежит под теменной долей, ограничивая с латеральной стороны центральную часть бокового желудочка. Хвост ядра участвует в образовании крыши нижнего рога бокового желудочка и достигает миндалевидного тела.
Наряду с полосатым телом в его составе выделяют стриатум, включающий хвостатое ядро и скорлупу. Стриатум и бледный шар образуют стриопаллидарную систему, которая, в свою очередь, относится к экстрапирамидной системе, участвующей в управлении движениями, регуляции мышечного тонуса.
Базальные ядра имеют сложные связи с корой полушарий большого мозга и с таламусом, через который они влияют на двигательные зоны коры. Базальные ядра участвуют в регуляции мышечного тонуса, управлении целенаправленными движениями, эмоциями и познавательными функциями.
Миндалевидное тело залегает в белом веществе височной доли полушария, примерно на 1,5-2 см кзади от височного полюса ниже скорлупы и кпереди от хвоста хвостатого ядра. Миндалевидное тело участвует в осуществлении оборонительного поведения, вегетативных, двигательных и эмоциональных реакций, является мотивацией условно-рефлекторных реакций.
Раздражение миндалевидного тела у человека ведет к изменению настроения (от гнева и страха до спокойствия и расслабления) в зависимости от эмоционального состояния непосредственно перед воздействием. Миндалевидное тело функционирует как «эмоциональный усилитель», его повреждения влияют на оценку эмоциональной окраски события.
Кроме серой коры на поверхности полушария, имеются еще скопления серого вещества в его толще, именуемые базальными ядрами и составляющие то, что для краткости называют подкоркой. В отличие от коры, имеющей строение экранных центров, подкорковые ядра имеют строение ядерных центров. Различают три скопления подкорковых ядер: corpus striatum, claustrum и corpus amygdaloideum.
1. Coprus striatum, полосатое тело, состоит из двух не вполне отделенных друг от друга частей — nucleus caudatus и nucleus lentiformis.
A. Nucleus caudatus, хвостатое ядро, лежит выше и медиальнее nucleus lentiformis, отделяясь от последнего прослойкой белого вещества, называемой внутренней капсулой, capsula interna. Утолщенная передняя часть хвостатого ядра, его головка, caput nuclei caudati, образует латеральную стенку переднего рога бокового желудочка, задний же утонченный отдел хвостатого ядра, corpus et cauda nuclei caudati, тянется назад по дну центральной части бокового желудочка; cauda заворачивается на верхнюю стенку нижнего рога. С медиальной стороны nucleus caudatus прилегает к таламусу, отделяясь от него полоской белого вещества, stria terminalis. Спереди и снизу головка хвостатого ядра доходит до substantia perforata anterior, где она соединяется с nucleus lentiformis (с частью последнего, называемой putamen). Кроме этого широкого соединения обоих ядер с вентральной стороны, имеются еще тонкие полоски серого вещества, располагающиеся вперемешку с белыми пучками внутренней капсулы. Они послужили причиной названия «полосатое тело», corpus striatum.
Редактор: Искандер Милевски. Дата последнего обновления публикации: 13.8.2020
Вентральный стриатум включает в себя ядро accumbens, вентральные части хвостатого ядра и putamen, и части обонятельного бугорка. Ядро accumbens разделено на периферически расположенную оболочку, окружающую центральную сердцевину, каждая из которых имеет разные типы клеток, соединения, синаптическую организацию и распределение рецепторов. Сердцевина ядра accumbens похоже на хвостатое ядро и putamen, в то время как оболочка демонстрирует более разнообразное нейрохимическое строение, а также более широкое разнообразие соединений. У приматов и грызунов оболочку и сердцевину можно отличить несколькими маркерами. Например, локализация кальбиндина и дофамина более плотная в ядре и разрежена в оболочке, тогда как локализация калретинина противоположна по своему характеру. Цитологически сердцевина и боковая оболочка содержат «колючие» нейроны среднего размера, которые меньше и несколько менее «колючие», чем те, которые встречаются на дорсальной части полосатого теле, в то время как медиальная оболочка содержит небольшие редко расположенные «колючие» нейроны. Как и дорсальный стриатум, ядро accumbens получает подавляющее большинство синапсов из коры. Дофаминергическая иннервация к ядру accumbens заканчивается гетерогенно с самым плотным представлением в медиальной оболочке.
В полосатом теле были идентифицированы два основных типа средних «колючих» нейронов, которые участвуют в двух классически трактов в базальных ганглиях, «прямой путь» и «косвенный путь». Важно отметить, что они не являются полностью независимыми и взаимосвязаны на нескольких уровнях. «Прямой путь» включает в себя гамкергические средние «колючие» нейроны, которые экспрессируют рецепторы P и дофамина D1. Эти нейроны посылают проекции на GPi и SNr и синапсы на гамкергические проекционные нейроны в обоих выходных ядрах. Таким образом, в довольно упрощенной схеме они препятствуют выходу гамкергических проекций из базальных ганглиев и, как следствие этого, ослабляют активность таламуса.
«Косвенный путь» включает в себя гамкергические средние «колючие» нейроны, которые экспрессируют энкефалин и имеют рецепторы дофамина D2. Эти нейроны контактируют с гамкергическими клетками GPe, которые проецируются в STN. Глутаматергические нейроны в проекциях STN отправляют свои проекции для GPi и SN. Когда средние «колючие» нейроны полосатого тела активируются глутаматергическими входами, они ингибируют GP, таким образом, обезвреживая глутаматергические нейроны STN. Усиленный возбуждающий сигнал к гамкергическим нейронам в GPi и SNr вызывает торможение таламуса.
Совсем недавно был описан третий путь базальных ганглиев, называемый «гиперректальным путем». Этот путь полностью избегает полосатого тела и состоит из серии связей между моторной корой, STN и GPi. Цель этого пути состоит в том, чтобы запретить уже начатые действия ( Aron and Poldrack, 2006 ). Исследования на животных и исследования изображений мозга показывают, что нарушение этого пути играет роль в импульсивном поведении.
Существует топографическая организация для дофаминергических связей, в которой проекции из наиболее дорзальных областей хвостатного ядра и скорлупы связаны с вентральными клеточными элементами мезенцефалических дофаминергических групп и наоборот. Нейроны на разных уровнях SNc имеют конкретные цели. Дорсальный уровень и медиальная часть проекций вентрального уровня к вентромедиальному стриатуму, который включает в себя ядро accumbens, части обонятельного бугорка и вентральные части хвостатного ядра и putamen. Остальная часть вентрального уровня выступает в центральные ассоциативные области полосатого тела, а столбцы дофаминергических клеток, которые простираются в проектции SNr до дорсолатеральной сенсомоторной зоны.
База́льные я́дра (также база́льные га́нглии, лат. nuclei basales) — несколько скоплений серого вещества, расположенных в белом веществе латеральнее таламуса на уровне основания полушарий конечного мозга. Базальные ядра входят в состав переднего мозга, расположенного на границе между лобными долями и над стволом мозга. Традиционно в состав базальных ядер включались полосатое тело (лат. corpus striatum), в свою очередь состоящее из хвостатого ядра (лат. nucleus caudatus), скорлупы (лат. putamen) и бледного шара (лат. globus pallidus), а также ограда (лат. claustrum) и миндалевидное тело (лат. corpus amygdaloideum). Бледный шар и скорлупа вместе называются чечевицеобразным ядром (лат. nucleus lentiformis). Белое вещество между таламусом и чечевицеобразным ядром называется внутренней капсулой (лат. capsula interna), между чечевицеобразным ядром и оградой — наружной капсулой (лат. capsula externa) и между оградой и островком — самой наружной капсулой (лат. capsula extrema). Эта классификация основана на топографии анатомического среза мозга, однако в последнее время она всё чаще заменяется функциональной, где под термином «базальные ядра» понимают полосатое тело и несколько ядер промежуточного и среднего мозга (субталамическое ядро (лат. nucleus subtalamicus), чёрная субстанция (лат. substantia nigra) и ножкомостовое ядро покрышки (лат. nucleus tegmentalis peduncolopontinus)), которые совместно обеспечивают функциональную регуляцию движений и мотивационных аспектов поведения. Функции ограды остаются недостаточно изученными, а структуры миндалевидного тела относят к лимбической системе.
а) Роль базальных ганглиев в когнитивном контроле двигательных актов. Контур хвостатого ядра. Понятие «когнитивная способность» означает мыслительные процессы мозга, использующие как сенсорную информацию, поступающую в мозг, так и информацию, уже накопленную в памяти. Большинство наших двигательных актов являются следствием реализации генерируемых в мозге мыслей — процесса, который называют когнитивным контролем двигательной активности. Главную роль в этом контроле играет хвостатое ядро.
Нервные связи между хвостатым ядром и кортикоспинальной моторной системой, показанные на рисунке ниже, слегка отличаются от связей контура скорлупы.
Контур хвостатого ядра системы базальных ганглиев для когнитивного планирования последовательных и параллельных двигательных программ для достижения специфических осознанных целей
Отчасти это связано с тем, что хвостатое ядро, как видно на рисунке ниже, распространяется во все доли большого мозга, начинаясь впереди в лобных долях, затем проходя через теменные и затылочные доли и, наконец, загибаясь снова вперед в височные доли, образуя подобие буквы «С».
Анатомические связи базальных ганглиев с корой большого мозга и таламусом, показанные в трехмерном изображении
Кроме того, хвостатое ядро получает основную часть своего «входа» из ассоциативных областей коры большого мозга, лежащих над хвостатым ядром, главным образом из областей, которые объединяют разные типы сенсорной и моторной информации в соответствующие мыслительные программы.
После того как сигналы из коры большого мозга достигают хвостатого ядра, они передаются к внутренней части бледного шара, затем к релейным ядрам переднего вентрального и вентролатерального таламуса и в итоге — назад к префронтальной, премоторной и дополнительной моторной областям коры большого мозга.
Однако практически ни один из возвращающихся сигналов не идет непосредственно к первичной моторной коре. Вместо этого сигналы идут к тем вспомогательным моторным регионам в премоторной и дополнительной моторной областях, которые имеют дело не с возбуждением отдельных мышечных движений, а с организацией программ последовательных движений, длящихся 5 сек или более.
Хорошей иллюстрацией сказанного будет мгновенная и автоматическая реакция человека, увидевшего приближение льва. Он:
(1) отворачивается от льва;
(3) даже пытается влезть на дерево.
Без когнитивных функций у человека не может быть интуитивного знания, позволяющего без слишком длительного размышления реагировать быстро и соответствующим образом.
Таким образом, когнитивный контроль двигательной активности определяет подсознательно и в течение секунд, какой комплекс движений будет полезен для достижения сложной цели, хотя само двигательное поведение в этом случае может продолжаться в течение многих секунд.
б) Участие базальных ганглиев в изменении временных характеристик и масштабного соотношения движений. В процессе регуляции движений реализуются две важные способности мозга:
(1) определять, как быстро движение должно выполняться;
(2) контролировать должную амплитуду движения.
Например, человек может писать букву «а» медленно или быстро. Так же он может написать маленькую «а» на листке бумаги или большую «а» на классной доске. Независимо от выбора пропорции буквы остаются примерно одинаковыми.
У больного с тяжелым поражением базальных ганглиев эти функции распределения во времени и масштабного соотношения движений недостаточны; иногда они практически отсутствуют. Базальные ганглии и здесь функционируют не в одиночку, а в тесной связи с корой большого мозга.
Особенно важной областью коры является задняя теменная кора, представляющая собой локус пространственных координат для регуляции движений всех частей тела, а также для соотношения тела и его частей со всем его окружением. На рисунке ниже показано, как человек, лишенный левой задней теменной коры, может рисовать лицо другого человека, изображая правильно пропорции с правой стороны, но почти игнорируя левую сторону (которая в его правом поле зрения).
Типичный рисунок, который может нарисовать человек с тяжелым поражением левой теменной коры, где хранятся пространственные координаты правого поля зрения
Такой человек также не будет пытаться пользоваться своей правой рукой, правой кистью или другими правыми частями тела для выполнения задач, почти не осознавая, что эти части его тела существуют.
Поскольку хвостатое ядро системы базальных ганглиев функционирует в основном в связи с ассоциативными областями коры большого мозга, например задней теменной корой, по-видимому, определение скоростных характеристик и масштабных соотношений движений является функциями этого контура когнитивного двигательного контроля хвостатого ядра.
Однако наше понимание функции базальных ганглиев все еще так неопределенно, что многое из того, что обсуждается в нескольких последних разделах, является предположением, логическим умозаключением, а не доказанным фактом.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
а) Прилежащее ядро. Полное название — прилежащее ядро прозрачной перегородки, «ядро, прислоненное к прозрачной перегородке». Точнее, это ядро примыкает к ядрам перегородки, расположенным в ее основании. Взаимоотношения этих структур можно увидеть на рисунках ниже. Прилежащее ядро — одна из глубоких областей мозга, которые пытались стимулировать при помощи электродов для облегчения болевого синдрома.
Стимуляция прилежащего ядра вызывает у испытуемого ощущение счастья (гедония), которое сравнимо с тем, которое испытывают люди после употребления наркотиков, например героина. Считают, что подобное чувство «эйфории» возникает из-за массивного синаптического выброса дофамина нейронами вентральной покрышечной области, который затем поступает к нейронам прилежащего ядра и медиальной префронтальной коры. В норме выброс дофамина происходит в небольшом количестве, после чего он быстро выводится из внеклеточного пространства за счет работы специфических транспортеров дофамина.
б) Область перегородки. Область перегородки состоит из ядер перегородки, которые спереди от передней комиссуры сливаются с корой больших полушарий. Ядра перегородки также частично продолжаются к самой прозрачной перегородке.
Ядра перегородки получают афферентные волокна из следующих источников: • от миндалевидного тела в составе тонкого диагонального пучка (Брока), который проходит вдоль переднего продырявленного вещества; • от обонятельного пути в составе медиальной обонятельной полоски; • от гиппокампа в составе свода; • от моноаминергических нейронов ствола мозга через медиальный пучок переднего мозга.
Эфферентные связи представлены двумя основными группами:
• Медуллярная полоска — пучок глутаматергических волокон, который идет вдоль места сочленения боковой стенки и крыши третьего желудочка. Эти волокна образуют синапсы с холинергическими волокнами ядра поводка. Ядра поводка правого и левого полушарий соединяет друг с другом спайка поводков, расположенная у корня эпифиза. От ядра поводка берет начало поводково-межножковый путь (возвратный пучок, пучок Мейнерта), который образует синапсы с нейронами межножкового ядра ретикулярной формации среднего мозга. Считают, что межножковое ядро вместе с нейронами голубого пятна принимает участие в регуляции цикла сон-бодрствование.
• Перегородочно-гиппокамповый путь, идущий в составе свода к гиппокампу. Его активностью объясняют появление на электроэнцефалограмме (ЭЭГ) медленноволнового гиппокампового в-ритма. Глутаматергичские нейроны этого пути служат водителями ритма, определяющими скорость θ-ритма; холинергические нейроны определяют размер 0-волн. θ-Ритм — следствие синхронного возбуждения пирамидных нейронов гиппокампа. Он чувствителен к биохимическим изменениям в структуре глутаматных рецепторов пирамидных нейронов, которые происходят в них при ДВП, необходимой для формирования отдельных воспоминаний.
У пациентов с болезнью Альцгеймера сила θ-ритма существенно снижена. Это связано с тем, что у таких больных происходит массивная гибель холинергических нейронов, страдает событийная память, нарушается процесс извлечения воспоминаний.
Связи области перегородки. ДМЯ—дорсальное медиальное ядро таламуса.Перегородочно-гиппокамповый путь (1) с обратными волокнами от гиппокампа (2).Сочленение среднего мозга и таламуса. П — правая сторона; Л — левая сторона.Часть поперечного среза через верхний отдел моста, показаны элементы ретикулярной формации.
в) Базальный отдел переднего мозга. Базальный отдел переднего мозга продолжается от бифуркации обонятельного пути кзади до уровня воронки гипофиза, от средней линии он продолжается до миндалевидного тела. На дне базального переднего мозга находится переднее продырявленное вещество, которое названо так потому, что через него проходят переднемедиальные центральные ветви, начинающиеся от виллизиевого круга. Здесь кора больших полушарий замещается рассеянными группами ядер, основное из которых — гигантоклеточное базальное ядро Мейнерта.
Тела холинергических нейронов расположены преимущественно в ядрах перегородки и базальном ядре Мейнерта. Базальное ядро отдает волокна ко всем участкам коры больших полушарий, которые также содержат рассеянные холинергические нейроны.
Перегородочные и базальные ядра вместе с небольшим числом ядер в диагональном пучке Брока часто объединяют под названием базальные ядра переднего мозга.
Во время бодрствования новая кора постоянно получает сигналы по холинергическим волокнам ядра Мейнерта, благодаря активности которого на ЭЭГ появляется характерный ритм «бодрствования». Все отделы новой коры богато иннервированы. Ацетилхолин (АХ) взаимодействует с мускариновыми рецепторами нейронов коры, снижает проницаемость их ионных каналов для калия и, таким образом, делает нейроны более восприимчивыми к другим возбуждающим стимулам. Холинергическая иннервация обеспечивает ДВП и синаптическое усиление пирамидных нейронов новой коры.
Общее замедление психических процессов, которое часто наблюдают у больных, перенесших инсульт, можно объяснить разрывом холинергических волокон в подкорковом белом веществе, причиной которого обычно бывает окклюзия передней или средней мозговой артерии. В результате нарушается холинергическая иннервация участков мозга, расположенных в зоне повреждения и сзади от нее.
Холинергическая иннервация базальных ядер переднего мозга корой больших полушарий. (А) Срез на уровне, показанном в (Б). (Б) Корковая иннервация. (В) Перегородочно-гиппокамповый путь в своде. Этот путь также иннервирует миндалевидное тело.
г) Нейрогенез и взрослый мозг. Нейрогенезом называют процесс формирования нейронов из стволовых клеток. В настоящее время хорошо известно, что нейрогенез происходит в течение всей взрослой жизни человека и (в меньшей степени) даже в пожилом возрасте. В головном мозге лабораторных животных (в том числе обезьян) и биопсийном материале, который забирают во время нейрохирургических операций, нейрональные стволовые клетки в стадии митоза были обнаружены в следующих двух зонах.
• В субвентрикулярной области, которая расположена сразу под эпендимальной выстилкой боковых желудочков. Отсюда происходят стволовые клетки обонятельных луковиц, которые описаны ранее. У взрослого человека из стволовых клеток субвентрикулярной зоны происходят клетки, которые в дальнейшем станут частью серого вещества лобной, теменной и височной долей, однако до настоящего времени неизвестно, трансформируются ли они в нейроны или нейроглию.
• В гиппокампальном комплексе имеется область, расположенная глубже слоя гранулярных клеток зубчатой извилины. У всех изученных видов, в том числе у человека, при исследовании культур стволовых клеток данной области было обнаружено, что эти клетки обладают электрической активностью и способны к ветвлению. Гистологические исследования, проведенные на крысах, позволили установить, что в дальнейшем эти стволовые клетки развиваются в полноценные зрелые гранулярные клетки.
У взрослых крыс число клеток, находящихся в процессе митоза, может резко возрастать при воздействии определенных чувствительных стимулов. Например, активность митоза в клетках обонятельных луковиц усиливается в пять раз, если крысу поместить в помещение с разнообразными запахами. Если же крысу заставляют обучаться чему-то новому (помещают в лабиринт, колесо и т.п.), возрастает интенсивность митоза в субгранулярной зубчатой зоне. Эти наблюдения еще больше убеждают ученых в том, что ментальные и физические упражнения благотворно сказываются на здоровье людей, приближающихся к пенсионному возрасту.
Благодаря экспериментам на животных существуют доказательства того, что лекарственные средства, применяемые для лечения нейродегенеративных и психических заболеваний, обладают положительным нейротрофическим эффектом. Повышение уровня серотонина во внеклеточном веществе зубчатой извилины стимулирует пролиферацию окружающих нейронов; данный эффект отмечают после введения ингибиторов обратного захвата серотонина или ингибиторов моноаминоксидазы.
1. Обонятельная система. Обонятельная система включает обонятельный эпителий полости носа, обонятельные нервы, обонятельные луковицы и обонятельные пути, а также несколько участков обонятельной коры. Обонятельный эпителий представлен биполярными нейронами, поддерживающими клетками и базальными клетками, которые отвечают за восстановление обонятельных нейронов в течение жизни (с возрастом скорость обновления нейронов снижается). Центральные отростки обонятельных нейронов формируют обонятельные нервы, которые проходят через ситовидную пластинку решетчатой кости и образуют синапсы с митральными клетками обонятельных луковиц.
Аксоны митральных клеток образуют обонятельный путь, который имеет окончания на нескольких участках передних отделов височной доли. За распознавание запахов отвечает орбитофронтальная кора, в которую информация поступает через дорсальное медиальное ядро таламуса.
2. Лимбическая система. Лимбическая система состоит из лимбической коры и связанных с ней подкорковых ядер. К лимбической коре относят кору гиппокампа, область перегородки, парагиппокампальную извилину и поясную извилину. Основное подкорковое ядро —миндалевидное тело. С лимбической системой тесно связаны орбитофронтальная кора, височный полюс, гипоталамус, ретикулярная формация и прилежащее ядро.
Передняя часть парагиппокампальной извилины представлена энторинальной корой, которая получает когнитивную и чувствительную информацию от ассоциативных участков коры. Далее она передает ее на гиппокампальный комплекс, где происходит консолидация информации. После этого информация передается обратно на ассоциативную кору, где происходит ее хранение в виде воспоминаний.
Гиппокампальный комплекс состоит из основания гиппокампа, собственно гиппокампа и зубчатой извилины. Отдельные секторы гиппокампа называют СА1-4.
Перфорантный путь идет от энторинальной коры до дендритов гранулярных клеток зубчатой извилины. Аксоны гранулярных клеток образуют синапсы с пирамидными нейронами САЗ, которые отдают коллатерали Шаффера к САГ СА1 проецируется обратно к энторинальной коре, которая плотно связана с ассоциативными участками коры.
При двустороннем повреждении или удалении гиппокампальной формации у пациента развивается антероградная амнезия, сопровождающаяся нарушением декларативной памяти. Процедурная память сохранена. Считают, что ключевую роль в консолидации воспоминаний играет долговременная потенциация гранулярных и пирамидных нейронов.
Функции островка связаны с языком (речью) и восприятием боли. Передняя поясная кора отвечает за выбор нужных двигательных реакций, эмоциональное состояние, тонус мочевого пузыря, вокализацию, а также контролирует вегетативную нервную систему. Задняя поясная кора определяет эмоциональные реакции, возникающие в ответ на зрительные или другие ощущения.
Миндалевидное тело, расположенное спереди и выше от височного рога бокового желудочка,—основное ядро головного мозга, отвечающее за чувство страха. Его афферентное (латеральное) ядро воспринимает обонятельные, зрительные, слуховые, тактильные и когнитивные стимулы, а также ощущения, исходящие от внутренних органов, и различные воспоминания. От эфферентного (центрального) ядра отходят нервные волокна, которые в составе терминальной полоски направляются к гипоталамусу. Они стимулируют выброс кортикотропина и вызывают вагус-опосредованную брадикардию.
От центрального ядра волокна направляются также к стволу мозга, где они возбуждают нейроны дорсального и вентрального околоводопроводного серого вещества, изменяют частоту дыхания и состояние вегетативной нервной системы. Амигдалофугальный путь, идущий от центрального ядра в составе лимбической петли полосатого тела, активирует поведение защиты/уклонения.
Область перегородки состоит из двух основных групп ядер. От одной группы в составе медуллярной полоски таламуса отходят глутаматергические волокна, направляющиеся к ядру поводка. От ядра поводка, в свою очередь, отходят холинергические волокна пучка Мейнерта, направляющиеся к межножковому ядру, регулирующему цикл сон-бодрствование. Другая группа ядер формирует перегородочно-гиппокамповый путь, который образует синапсы с пирамидными нейронами гиппокампа. Глутаматергические и холинергические нейроны определяют (соответственно) скорость и выраженность гиппокампового 0-ритма, от которого зависит формирование эпизодических воспоминаний.
Базальный отдел переднего мозга — участок серого вещества вокруг переднего продырявленного вещества и в нем самом. Здесь расположено холинергическое базальное ядро Мейнерта, которое отдает волокна ко всем участкам коры больших полушарий. Здесь также находится холинергическое ядро перегородки, проецирующееся к гиппокампу. Примерно половина этих нейронов погибает при болезни Альцгеймера, а пути, которые соединяют эти клетки с корой больших полушарий, могут повреждаться при инсульте.
Редактор: Искандер Милевски. Дата публикации: 25.11.2018







 Контур хвостатого ядра системы базальных ганглиев для когнитивного планирования последовательных и параллельных двигательных программ для достижения специфических осознанных целей
Контур хвостатого ядра системы базальных ганглиев для когнитивного планирования последовательных и параллельных двигательных программ для достижения специфических осознанных целей Анатомические связи базальных ганглиев с корой большого мозга и таламусом, показанные в трехмерном изображении
Анатомические связи базальных ганглиев с корой большого мозга и таламусом, показанные в трехмерном изображении Типичный рисунок, который может нарисовать человек с тяжелым поражением левой теменной коры, где хранятся пространственные координаты правого поля зрения
Типичный рисунок, который может нарисовать человек с тяжелым поражением левой теменной коры, где хранятся пространственные координаты правого поля зрения Связи области перегородки. ДМЯ—дорсальное медиальное ядро таламуса.
Связи области перегородки. ДМЯ—дорсальное медиальное ядро таламуса.  Перегородочно-гиппокамповый путь (1) с обратными волокнами от гиппокампа (2).
Перегородочно-гиппокамповый путь (1) с обратными волокнами от гиппокампа (2).  Сочленение среднего мозга и таламуса.
Сочленение среднего мозга и таламуса.  Часть поперечного среза через верхний отдел моста, показаны элементы ретикулярной формации.
Часть поперечного среза через верхний отдел моста, показаны элементы ретикулярной формации.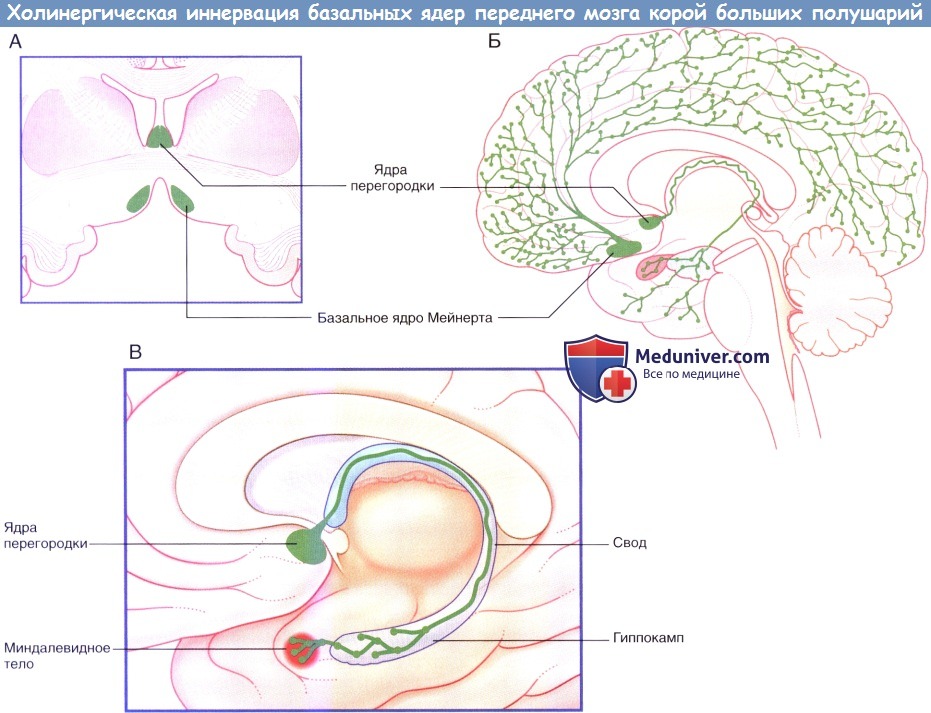 Холинергическая иннервация базальных ядер переднего мозга корой больших полушарий.
Холинергическая иннервация базальных ядер переднего мозга корой больших полушарий.